Vantagens da pecuária brasileira X imagem dos concorrentes
21 de fevereiro de 2003Comportamento do sistema radicular do capim Tanzânia irrigado
21 de fevereiro de 2003Efeitos da monensina sobre as bactérias ruminais em dietas ricas em volumoso ou concentrado
O ionóforo monensina vem sendo utilizado há muitos anos na bovinocultura de corte com intuito de melhorar a eficiência de utilização dos subprodutos da fermentação ruminal, através da diminuição da produção de metano e aumento do ácido propiônico. A monensina atua sobre a microbiota ruminal diminuindo a concentração de bactérias gram positivas, produtoras de metano. Com a queda da produção de metano, os produtos resultantes da fermentação de carboidratos são utilizados na produção de propionato, evitando desperdício de energia na formação de gases. Porém, resultados obtidos por Hungate (1996), citado por Lana & Russell, (2001) mostram que bactérias obtidas diretamente do rúmen de animais recebendo monensina, apresentaram coloração gram variável.
Em outra pesquisa, Dawson & Bowling (1983) observaram que bovinos tratados com monensina possuíam uma população bacteriana ruminal mais tolerante a monensina in vitro que a população bacteriana de animais controle. A monesina atua sobre as bactérias sensíveis, catalisando a saída de K. Quando bovinos receberam 350 mg de monensina por dia, a concentração de monensina necessária para causar metade da perda máxima de K aumentou 8 vezes, e essa mudança ocorreu dois dias após o início da suplementação (Lana & Russell, 2001).
A utilização de monensina para animais confinados recebendo grandes quantidades de cereais, causou um aumento na eficiência alimentar de 6,4% (resultados de 228 ensaios). Já na utilização para animais em pastejo, foi observada melhora de 13,5% em ganho de peso para animais suplementados em relação ao controle (Goodrich et al., 1984). Esses valores sugerem que bactérias ruminais de animais em pastagem, são mais sensíveis a monensina do que bactérias de animais recebendo grandes quantidades de concentrado.
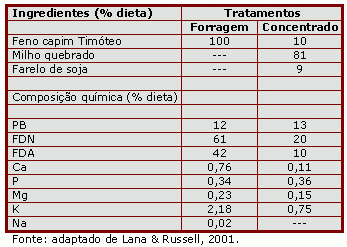
Lana & Russel (2001) compararam os efeitos da monensina sobre os produtos da fermentação, e a resistência bacteriana à perda de K intracelular, com aumento gradativo do ionóforo in vitro, em microorganismos provenientes de animais recebendo forragem ou dieta rica em concentrado. Essas técnicas permitem acessar a sensibilidade das bactérias ruminais aos ionóforos, nos quais maior sensibilidade em função da dieta é indicativo de maior potencial de mudança na população microbiana ruminal pelo uso de ionóforos, com maiores benefícios ao desempenho animal. Foram utilizados 4 bovinos fistulados no rúmen, alimentados com 100% de forragem ou 90% de concentrado e 10% de forragem, limitando o consumo a 95% do consumo ad libitum. Foi utilizado feno de capim Timóteo como volumoso, milho quebrado e farelo de soja na composição das dietas, como descrito na tabela 1. Água e sal mineral foram fornecidos à vontade.
Tabela 1. Composição das dietas experimentais.
Foi coletada digesta de vários pontos do rúmen para avaliação de pH, ácidos graxos voláteis (AGV) e isolamento de bactérias. No experimento para avaliação da resistência das bactérias a perda de potássio intracelular em resposta ao uso crescente de monensina, bactérias ruminais foram transferidas anaerobicamente para tubos contendo de 0 a 6,4 mM de monensina e incubados à 39oC. O K foi quantificado em espectofotômetro de chama, sendo a perda de K estimada pelo decréscimo de K intracelular. Foram obtidas equações de regressão linear da perda de K com a concentração de monensina, determinando-se a quantidade de monensina necessária para causar a metade da perda máxima de potássio intracelular (Kd) e a perda máxima potássio intracelular (Kmax).
Animais recebendo dietas com altos níveis de concentrado apresentaram pH variando de 6,2, imediatamente antes da alimentação, a 5,5, 3 horas após a alimentação. Para esses mesmos animais, a concentração de AGV foi de 130 mM e a relação acetato:propionato de 1,5:1. Na dieta contento somente volumoso, o pH ruminal foi mais constante, aproximadamente 6,7, e a concentração de AGV de 80 mM, sendo a relação acetato:propionato 4:1.
Quando as bactérias provenientes de dietas com altos níveis de concentrado ou de dietas exclusivas de forragens foram incubadas com feno de gramíneas por 24 h, a produção total de AGV foi de 64 e 72 mM, respectivamente. As bactérias provenientes de dieta rica em concentrado, produziram menos acetato, mais propionato e menos metano, que bactérias provenientes da dieta exclusiva de volumoso, sendo as taxas de acetato:propionato 1,75:1 e 3,75:1, respectivamente. A menor produção de metano das bactérias provenientes da dieta com alto teor de concentrado foi provavelmente devido a menor população de microorganismos metanogênicos existentes in vitro, assim como in vivo, devido à sua inibição pelo pH ruminal. A monensina acarretou pequeno, mas significante (P<0,001) decréscimo na produção de AGV e reduzida relação acetato:propionato. As bactérias de animais que receberam forragem foram mais sensíveis à monensina que as bactérias da dieta com altos teores de concentrado. Quando a monensina foi aumentada de 0 a 10 uM (micromolar), a relação acetato:propionato diminuiu de 3,8 para 2,4 com bactérias de animais com 90% de concentrado, e somente 0,5 (1,75 para 1,25) para bactérias de animais alimentados exclusivamente com forragem. Para bactérias provenientes de animais com dieta à base de forragem, pequenas concentrações de monensina já causaram queda na produção de metano, continuando a diminuir com o aumento das concentrações. Para bactérias com dieta de 90% concentrado, não houve resposta para redução de metano até a concentração de monensina alcançar 0,5 uM. Os valores de Kmax das bactérias ruminais da dieta com 90% de concentrado ou exclusivas de forragem foram semelhantes, mas o Kd das bactérias com dieta de 90% concentrado foi 12 vezes maior que das bactérias provenientes de dietas à base de volumoso. Esses resultados mostram que as bactérias de animais com dietas exclusivas de forragem são mais sensíveis à monensina do que bactérias oriundas de animais com altos teores de concentrado. A monensina sempre diminuiu a produção de metano e a relação acetato:propionato, mas a resposta em função do aumento do nível de monensina foi maior nas bactérias de animais alimentados com forragem. Referências bibliográficas
DAWSON, K.A.; BOLING, J.A. Monensin resistant bacteria in the rumen of calves on monensin containing and unmedicated diets. Appl. Envirom. Microbiolog., v.46, p. 160-164, 1983.
GOODRICH, R.D.; GARRET, J.E.; GAST, D.R. Influence of monensin on the performance o cattle. J. Anim. Sci., v.58, p. 1484-1498, 1984
LANA, R.P.; RUSSELL, J.B. Efeitos da monensina sobre a fermentação e sensibilidade de bactérias ruminais de bovinos sob dietas ricas em volumos ou concentrado. Rev. Bras. Zoot., v. 30, p. 254-260, 2001.
