Angus irá fazer abate teste no Bertin Lins/SP
12 de julho de 2005MT: leilões da Expoagro têm preços mais baixos esse ano
13 de julho de 2005Reprodução programada de touros jovens: retrospectiva
Por Prof. Dr. Arcadio de los Reyes Borjas1
No atual ano 2005 o Programa de Melhoramento genético da Raça Nelore – USP (PMGRN) comemora 10 anos da publicação do primeiro “Sumário de Avaliação Genética de Touros, Matrizes e Animais Jovens” (Lôbo et al., 1995), através de predições BLUP com a metodologia de Modelos Mistos sob Modelo Animal, marcando o início de uma grande expansão no uso dessa metodologia em gado de corte no Brasil.
Esse fato foi precedido por uma intensificação do trabalho da equipe técnica, criadores participantes e demais colaboradores do PMGRN, iniciada no segundo semestre de 1993, com o objetivo de aprimorar o sistema de controle e manutenção da base de dados e os restantes componentes da infra-estrutura, tais como recursos computacionais e pessoal técnico e de apoio necessários, incluindo a posta em funcionamento do recém lançado, nesse ano, aplicativo MTDFREML para a estimativa de (co)variâncias e predições BLUP de valores genéticos sob modelos mistos (Boldman et al., 1993).
Como resultado desse trabalho, em 1994 foi obtida a primeira avaliação genética sobre a base de dados do PMGRN sob modelo animal, e embora essa avaliação não tenha sido oficialmente publicada, os modelos matemáticos testados e as estimativas de (co)variâncias e parâmetros genéticos obtidas constituíram a base para o trabalho posterior. Tais estimativas foram apresentadas no “5th World Congress on Genetics Applied to Livestock Production” no Canadá (Lôbo et al., 1994).
O anterior propiciou um excelente ambiente de trabalho para o surgimento e debate de idéias e iniciativas entre especialistas, técnicos, alunos da pós-graduação e criadores participantes, sobre quais ações poderiam contribuir para o aprimoramento do PMGRN.
Nesse contexto criou-se o Centro de Avaliação de Tourinhos (CAT), que funcionou durante algum tempo no Rancho Guanacaste, Uberaba-MG. Na procura para aperfeiçoar o processo de avaliação e seleção de touros jovens, como elo determinante principal do progresso genético a ser alcançado, idealizamos e propusemos a implantação da Reprodução Programada (RP) de machos jovens, a qual teve inicio em 1995, à par do lançamento do primeiro Sumário de Avaliação Genética do PMGRN.
Em outubro de 1995 elaboramos o documento base inicial contendo os princípios e objetivos da RP, para sua divulgação entre os participantes do PMGRN (Reyes, 1995). Posteriormente foi publicado um segundo artigo com argumentos mais completos sobre o papel da RP na consecução do progresso genético (Reyes et al., 1997), apresentado no 5o Seminário do PMGRN, Ribeirão Preto-SP, o qual, pela sua vigência, inclui-se na íntegra em ANEXO.
Como acontece logicamente com a implantação de qualquer novo processo, que implique mudanças do tradicional ou estabelecido, a RP enfrentou inicialmente certa resistência ou foi aceita com reservas e dúvidas, no entanto, os resultados positivos, que começaram a ser palpáveis de maneira relativamente rápida, determinaram uma adesão crescente por parte dos criadores participantes, que a partir de 1998 ultrapassou amplamente a oferta possível de sêmen dos touros incluídos na RP a cada ano, assim, o número de touros tem aumentado sistematicamente, quase quintuplicando entre 1998 e 2003, período em que o número de rebanhos participantes da RP quase quadruplicou (Marcondes et al., 2004). Dessa maneira, a RP tem-se consagrado como componente diferencial do PMGRN.
A RP do PMGRN está completando 10 anos em 2005, comemorando resultados como os apresentados em publicações recentes (Nassir et al., 2002; Bezerra, 2003; Marcondes et al., 2004). No entanto, é imperativo se questionar em que medida nesse período foram atingidos os objetivos inicialmente propostos (ANEXO), e quanto se tem avançado no aprimoramento do processo para liberar sua potencialidade máxima, e identificar se existem fatores dentro do próprio processo da RP ou, mais importante ainda, no seu papel na materialização do progresso genético de toda a população, que merecem ser reformulados e/ou aperfeiçoados.
Nesse último aspecto resulta importante observar, segundo expressa a equação (1) de predição do progresso genético apresentada no ANEXO, que a taxa do progresso genético por unidade de tempo, para uma característica e população determinada, depende de maximizar o produto AC x IS (acurácia da avaliação x intensidade de seleção), e minimizar o IG (intervalo de gerações), dado que a variabilidade genética (sA) pode-se considerar constante para os propósitos do cálculo, assim, o entendimento das inter-relações dessas variáveis com todo o processo de avaliação genética, seleção e acasalamentos permite a criadores e técnicos tomar as decisões mais adequadas, especialmente na seleção e uso dos touros.
Os objetivos da RP, definidos no ANEXO, visam maximizar o progresso genético através do elo reconhecidamente mais importante do melhoramento genético em bovinos: a avaliação, seleção e uso dos touros. No entanto, para atingir esses objetivos, não basta dispor da metodologia de avaliação genética mais acurada e aplicar a máxima intensidade de seleção possível, com estrito controle sobre a consangüinidade no meio e longo prazo, é também indispensável manter o intervalo de gerações tão reduzido quanto possível.
Aos efeitos da taxa de progresso genético que pode ser alcançada, o conceito de intervalo de gerações não se refere apenas ao fato dos reprodutores serem sexualmente mais precoces e atingirem a puberdade à menor idade, pois, do ponto de vista do melhoramento genético, o IG é uma função da idade dos indivíduos utilizados como reprodutores quando nascem suas progênies. Dessa forma o IG, num momento determinado, expressa a idade média dos pais ponderada pelo número de descendentes produzidos.
Fica claro que a prática comum de usar intensamente os touros mais velhos contribui expressivamente para o aumento do IG, reduzindo, em conseqüência, a taxa anual do progresso genético alcançável, sempre que se disponha de outros mais jovens com igual ou superior potencial genético.
Com base nas idéias prévias é simples deduzir que os reprodutores (vacas e touros), a medida que envelhecem, teriam que dar uma contribuição maior ao progresso genético através do valor genético médio de suas progênies para contrabalançar a redução devida ao aumento do IG, mas, isto é impossível, já que o valor genético real (genótipo) de um individuo não muda durante toda sua vida, enquanto a predição de tal valor genético (ex. DEP) é uma magnitude relativa, expressa como desvio de um certo valor genético médio da população avaliada, a base genética adotada.
Assim sendo, se existe progresso genético (mudança positiva ou negativa, segundo a natureza de cada característica, na média genética da população) a predição do valor genético de qualquer individuo expressará a cada ano uma mudança desfavorável na sua ordem de mérito respeito da base genética, ficando claro que o processo de contrabalanceamento é biologicamente impossível.
O raciocínio anterior sugere que a taxa de substituição dos touros na reprodução, especialmente sob inseminação artificial (IA), deveria ser a máxima possível, o qual implica reduzir ou eliminar o uso dos reprodutores mais velhos para viabilizar a difusão do material genético superior dos reprodutores mais jovens.
A prática atual está, em grande medida, na contramão desse princípio, sendo comum nas Centrais de IA a existência e comercialização de sêmen de touros com mais de 20 anos, e touros com mais de 15 anos ainda produzindo sêmen, sendo que todos eles já têm milhares de filhos na população, a exemplo do que acontece na raça Nelore.
Isto constitui a principal causa determinante do aumento do parentesco médio (co-ancestralidade) na população de uma raça, com o conseqüente incremento da consangüinidade, com taxa crescente a cada geração, e redução inevitável do progresso genético, além da devida ao maior IG, a ocasionada pela significativa perda por depressão consangüínea, cujos conhecidos efeitos prejudicais são mais expressivos sobre as características de maior impacto sobre o retorno econômico dos rebanhos de corte, aquelas ligadas à capacidade adaptativa dos animais, como saúde, habilidade maternal, eficiência reprodutiva e capacidade de sobrevivência (Burrow, 1993; McDaniel, 2001; Parnell, 2002).
Isto permite estabelecer que, se tratando de características de interesse econômico para a produção bovina, “a melhor maneira de se usar a consangüinidade é evitá-la”.
Caberia se questionar, se ao invés de comemorar cada aniversário desses “monstros sagrados da raça”, ou seus milhares de doses de sêmen estocados ou em processo de produção, dever-se-ia sentir um profundo constrangimento por não ter conseguido substituir qualquer um deles por outro mais jovem e superior geneticamente, dentre aqueles das seguintes gerações da mesma família (linhagem), ou muito melhor ainda, de outra linhagem ou produto de acasalamento dirigido entre as restantes que compõem a população da raça.
Vale ressaltar que não se trata da substituição física de tal ou qual reprodutor, mas sim, da exclusão real de seu material genético da reprodução provedora de reprodutores (machos e fêmeas) dos rebanhos geradores e multiplicadores do progresso genético: a população ativa.
A seleção com base nas predições BLUP dos valores genéticos, e a aplicação de biotecnologias na reprodução de fêmeas, têm incrementado a taxa de progresso genético alcançável, mas também tem determinado uma taxa crescente de consangüinidade, reduzindo a resposta à seleção e a variabilidade genética no médio e longo prazo, devido à amostragem Mendeliana entre as progênies de um conjunto particular de progenitores.
Esse processo conduz à acumulação da co-ancestralidade e consangüinidade, via o enorme impacto de poucos indivíduos ou famílias selecionados, em conseqüência da maior probabilidade de co-seleção de animais aparentados (Wiggans et al., 1995; Sánchez et al., 1999; VanRaden e Smith, 1999; Muir, 2000; Bijma et al., 2001; Weigel, 2001; Weigel e Lin, 2002).
Schenkel et al. (2002) estimaram a consangüinidade e os efeitos da depressão consangüínea individual e materna (vacas) nas raças Gir, Guzerá, Indubrasil, Nelore e Tabapuã, para o ganho médio diário pré-desmama (GMD205) e pós-desmama (GMD550).
Exceto na raça Guzerá, a taxa anual de consangüinidade foi crescente, particularmente após 1980, sendo que a proporção de animais consangüíneos aumentou rapidamente ao longo dos anos, tendo chegado, em 1999, a 60 % nas raças Nelore e Indubrasil e superior a 30% nas demais. A depressão consangüínea, por cada 10 % de aumento na consangüinidade individual, foi de 1,7 % (GMD205) e 2,1 % (GMD550) relativo à média fenotípica, com resultados correspondentes para a consangüinidade materna de 1,4 % e 0,5 %.
No Sumário 2003 do PMGRN (Lôbo et al., 2003) foram incluídos 68 touros participantes da RP, com idades, nessa altura, entre 5 e 15 anos, sendo que 17 deles (25 %) tinham 10 anos ou mais, 29 (42,6 %) 7 a 9 anos e 22 (32,4 %) entre 5 e 6 anos. Esses 68 touros são filhos de 39 pais, existindo entre os mesmos duas famílias Pai-Filho (Pai: G 2544/Filhos-I 3360, ZAN J 0039; Pai: F 8200/Filho-I 9940).
Uma análise da genealogia disponível dos 68 touros pela linha paterna, incluindo até a sexta geração, revelou que são descendentes de sete ancestrais, cinco Nelore Padrão (RG: 2822, 2850, 3987, 6720, A 0027) e dois Nelore Mocho (H 0064, H 4018).
O anterior, ainda sem considerar a genealogia pelo lado materno, implica um alto grau de parentesco (co-ancestralidade) de grande número dos 68 touros, em particular levando-se em conta que 57 (83,8 %) deles pertencem às linhagens paternais de três genearcas (2822, 3987 e H 0064). Esses resultados estão em correspondência com os achados por Vozzi e Lôbo (2004).
Usando o Mérito Genético Total (MGT) como critério de seleção, além de outras restrições, os 68 touros estão incluídos no percentil 30% da população para esse índice.
Por outro lado, incluem-se 21 touros (30,9%) que não atingem o percentil 30% (0,08) para a DPE450 (DEP para Perímetro Escrotal (PE) aos 450 dias de idade), em quanto 17 (25%) não classificam no percentil 50 % (0,41) para MP120 (DEP maternal para peso aos 120 dias de idade).
Sendo o PE o critério de seleção integrado no MGT para a eficiência reprodutiva, e levando em conta a reconhecida importância da eficiência reprodutiva e da habilidade maternal no retorno econômico da pecuária bovina, valeria a pena re-considerar os critérios atualmente aplicados.
O Sumário eletrônico 2004 do PMGRN da raça Nelore apresenta o Catálogo de Touros da RP, com 89 touros incluídos no percentil 25% para MGT, mesmo com maior número e algumas mudanças individuais, a estrutura de idades, ascendência genealógica e caracterização genética geral do grupo são similares às descritas previamente para o grupo da RP 2003.
O Sumário eletrônico 2004 do PMGRN da raça Nelore, apresenta 57 touros aprovados TOP 25% para MGT, nascidos entre 1976 e 1995 (idades entre 9 e 28 anos), deles 19 (33,3%) com 15 anos ou mais, 32 (56,2%) entre 10 e 14 anos e 6 (10,5%) com 9 anos. Dos 57 touros, 16 (28,1%) não classificam no percentil 50% (-0,03) para DPE450, e ainda também 16 não entram no percentil 50% (0,41) para MP120, sendo que 4 (7%) fazem parte de ambos subconjuntos.
Os critérios primários de admissão de machos jovens na RP têm-se extendido a considerações diferentes da avaliação genética efetuada pelo PMGRN, com a inclusão de animais procedentes de Provas de Desempenho Individual (PDI) e outros, a exemplo da PDI que desenvolve o CNPAF-EMBRAPA (DBO Sul-Revista Nelore 097, 2004), parte de cujos animais procedem de rebanhos do próprio PMGRN.
A avaliação numa PDI pode atingir uma acurácia máxima h (raiz quadrada da herdabilidade do caráter usado como critério), muito inferior à alcançada através do BLUP (Modelo Misto). Isto seria como tentar conferir os resultados obtidos através de programas e computadores modernos usando-se um ÁBACO. Processo que acrescenta gastos desnecessários, de tempo e recursos, à avaliação do animal.
O anterior não significa que não sejam utilizados critérios complementares de interesse econômico (conformação, raciais, etc.), mas que os mesmos sejam ponderadamente aplicados dentro do grupo de machos pré-selecionados pelo valor genético agregado. Essa flexibilização desqualifica, em grande medida, os objetivos e critérios inicialmente preconizados, e poderia comprometer a contribuição esperada da RP ao progresso genético da população, por cuja razão valeria a pena reconsiderar a mesma.
Os problemas expostos previamente afetam, não apenas à RP e aos rebanhos participantes do PMGRN-USP, mas sim a raça Nelore como população, e muito provavelmente a grande parte das restantes populações zebuinas de bovinos de corte no Brasil.
Referências bibliográficas
Bezerra, L.A.F. Reprodução Programada – Quem acredita, ganha!. 2003. Disponível em: http://www.ancp. org.br/reproducao/001.htm. Acessado em 28 de setembro de 2004.
Bijma, P.; Van Arendonk, J.A.M.; Woolliams, J.A. Predicting rates of inbreeding for livestock improvement schemes. J. Anim. Sci., 79: 840-853, 2001.
Boldman, K.G.; Kriese, L.A.; Van Vleck, L.D. and Kachman, S.D. 1993. A manual for use of MTDFREML. A Set of Programs to Estimates of Variances and Covariances. USDA, ARS, USA.
Burrow, H.M. The effects of inbreeding in beef cattle. Anim. Breed. Abstr., 61(11): 737-751, 1993.
Catálogo de Touros da RP 2004. Disponível em: http://www.ancp.org.br/sumarios/ catalogo.asp. Acessado em 31 de outubro de 2004.
DBO Sul-Revista Nelore Ed. 097, 2004. Disponível em: http://www.dbosul.com.br /revistas/revista_Nelore/pdbos_Nelore.asp?Edicao=097. Acessado em 25 de abril de 2005.
Lôbo, R.B.; Bezerra, L.A.F.; Oliveira, H.N.; Magnabosco, C. de U.; Zambianchi, A.R.; Albuquerque, L.G.; Bergmann, J.A.G.; Sainz, R.D. Avaliação Genética de Animais Jovens, Touros e Matrizes. Ribeirão Preto, GEMAC -Departamento de Genética – FMRP – USP. 94 p., 2003.
Lôbo, R.B.; Reyes, A. de los; Bezerra, L.A.F. Avaliação Genética de Touros, Matrizes e Animais Jovens – Sumário 1995, 67+VI p. 1995.
Lôbo, R.B.; Reyes, A. de los; Ferraz, J.B.S.; Bezerra, L.A.F.; Mercadante, M.E.Z.; Salles, P.A. Bivariate Animal Model analysis of growth weights and scrotal circumference of Nellore cattle in Brazil. Proc. 5th World Congress on Genetics Applied to Livestock Production, 17: 199-201, University of Guelph, Ontario, Canada, 1994.
Marcondes, C.R.; Zambianchi, A.R.; Freitas, M.A.R.; Lôbo, R.B. 2004. Perfil de escolha pelos touros da Reprodução Programada do PMGRN-Nelore Brasil do ano de 2003. V Simpósio da Sociedade Brasileira de Melhoramento Animal, SBMA, 2004. Disponível em: http://www.sbmaonline.org.br/ anais/v/bc040.pdf. Acessado em 30 de outubro de 2004.
McDaniel, B.T. Uncontrolled inbreeding. J. Dairy Sci., 84 (E. Suppl.): E185-E186, 2001.
Muir, W.M. The interaction of selection intensity, inbreeding depression, and random genetic drift on short- and long-term response to selection: Results using finite locus and finite population size models incorporating directional dominance. Proc. of the American Society of Animal Science, 2000.
Nassir, L.C.; Freitas, M.A.R.; Siqueira, R.L.P.G.; Figueiredo, L.F.C.; Cardoso, M.C.P.; Marcondes, C.R.; Maffei, W.E.; Lôbo, R.B. Reprodução Programada como acelerador do progresso genético nos rebanhos do NELORE BRASIL. IV Simpósio da Sociedade Brasileira de Melhoramento Animal, SBMA, 2002. Disponível: http://www.sbmaonline.org.br/anais/iv/ivt14bc.pdf. Acessado em 30 de outubro de 2004.
Parnell, P. Pedigree analysis of the Australian Angus population. Paper presented at Angus Society of Australian Workshop “Breeding and Management to meet Demand”, 2nd October, 2002, Wagga Wagga, NSW.
Reyes, A. de los. Por que e para que reproduzir o material genético de machos jovens no PMGRN. Material elaborado para divulgação entre os participantes do PMGRN, outubro de 1995.
Reyes, A. de los; Trovo, J.B.F.; Lôbo, R.B. Reprodução programada de machos jovens: acelerador do progresso genético. Documento técnico apresentado no 5o Seminário do PMGRN. Ribeirão Preto, 11 de abril de 1997.
Sánchez, L.; Toro, M.A.; García, C. Improving the efficiency of artificial selection: More selection pressure with less inbreeding. Genetics, 151: 1103-1114, 1999.
Schenkel, F.S.; LaGioia, D.R.; Riboldi, J. Níveis de endogamia e depressão endogâmica no ganho de peso de raças Zebuínas no Brasil. IV Simpósio da Sociedade Brasileira de Melhoramento Animal, SBMA, 2002. Disponível em: http://www.sbmaonline.org.br/anais/iv/ivt08bc.pdf. Acessado em 29 de outubro de 2004.
VanRaden, P.M. e Smith, L.A. Selection and mating considering expected inbreeding of future progeny. J. Dairy Sci., 82: 2771-2778, 1999.
Vozzi, P.A. e Lôbo, R.B. Variabilidade genética nos rebanhos e na reprodução programada do PMGRN. 2004. Disponível em: https://beefpoint.com.br /bn/radarestecnicos/default.asp?area=14&area_desc=Melhoramento+Genético. Acessado em 28 de outubro de 2004.
Weigel, K.A. Controlling inbreeding in Modern Breeding Programs. J. Dairy Sci., 84(E. Suppl.): E177-E184, 2001.
Weigel, K.A. and Lin, S.W. Controlling inbreeding by constraining the average relationship between parents of young bulls entering AI Progeny Test Programs. J. Dairy Sci., 85: 2376-2383, 2002.
Wiggans, G.R.; VanRaden, P.M.; Zuurbier, J. Calculation and use of inbreeding coefficients for genetic evaluation of United States dairy cattle. J. Dairy Sci., 78: 1584-1590, 1995.
ANEXO
Reprodução programada de machos jovens: acelerador do progresso genético
Documento técnico para o Seminário do PMGRN realizado em abril de 1997, elaborado pelos Prof. Dr. Arcadio de los Reyes Borjas, Pesquisador Visitante – Bolsista RHAE-CNPq, Dr. José Benedito de Freitas Trovo, Pesquisador EMBRAPA-CENARGEN-MA e Prof. Dr. Raysildo Barbosa Lôbo, Coordenador do PMGRN
A eficiência de um programa de melhoramento, medida como o progresso genético alcançado por unidade de tempo, depende da acurácia da avaliação genética dos candidatos a progenitores (pais) da próxima geração, da intensidade de seleção praticada (determinada pelo número de candidatos avaliados) e da magnitude do intervalo de gerações (tempo médio desde o nascimento dos reprodutores em uso até que estes são substituídos por outros jovens), isto representa a idade média dos reprodutores que estão sendo utilizados.
Assim, a magnitude do progresso genético anual é expressa pela seguinte equação:
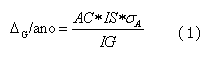
onde,
DG/ano = taxa de ganho genético anual;
AC = acurácia media na predição do valor genético dos candidatos á seleção;
IS = intensidade de seleção aplicada, depende da proporção selecionada;
IG = intervalo de gerações;
sA = desvio padrão genético, medida da variabilidade na população.
Da análise dos elementos da equação (1), fica claro que o máximo ganho genético anual depende de maximizar o produto AC x IS, e minimizar o IG, dado que a variabilidade genética (sA) é constante para os propósitos do cálculo, assim, o entendimento das relações entre estas variáveis, permite a criadores e técnicos tomar as decisões mais adequadas em termos da seleção.
Uma valorização exagerada da acurácia, como erradamente têm acontecido muitas vezes, pode prejudicar a taxa de ganho genético anual nos rebanhos, já que se, por exemplo, na hora de selecionar os reprodutores a serem usados, restringe-se o número de candidatos, escolhendo-se apenas entre aqueles com a maior acurácia, a intensidade de seleção será menor que quando aceitam-se níveis moderados da mesma.
Note-se que os touros com mais alta acurácia têm grande número de progênies avaliadas, sendo que, se existe progresso genético, é altamente provável encontrar filhos ou até netos desses touros com maior mérito genético do que seus progenitores, mesmo que as acurácias sejam menores. Na prática, decisões baseadas na acurácia podem ter pouco ou nenhum efeito sobre o progresso genético dos rebanhos, especialmente sob condições de avaliação com Modelo Animal, usando-se todas as informações disponíveis.
Dessa forma, verifica-se que a eficiência de um programa sustenta-se na avaliação genética mais acurada possível e à menor idade dos candidatos à seleção e, conseqüentemente, na disponibilidade do maior número de animais jovens avaliados a fim de poder aplicar a máxima intensidade de seleção.
Em gado de corte, como em outras populações animais, especialmente sob Inseminação Artificial (IA), o papel do macho reprodutor é decisivo em função do grande número de progênies que este pode produzir. Assim sendo, sua seleção e uso têm que seguir critérios rigorosos quando se quer garantir a multiplicação em grande escala do material genético efetivamente superior.
Durante muito tempo a avaliação genética de machos reprodutores em gado de corte foi efetuada, quase exclusivamente, por meio de provas de desempenho individual de animais jovens (Provas de Ganho em Peso). Nos últimos, anos têm ocorrido um enorme avanço nos métodos de predição do valor genético usando dados de campo o que, junto ao astronômico progresso da computação e ao uso crescente da IA em gado de corte, tem criado uma estrutura de informações cada vez mais adequada para a aplicação de metodologias avançadas na avaliação genética.
É necessário ressaltar que não basta dispor de recursos computacionais potentes e sistemas de avaliação genética altamente sofisticados. Criadores e melhoristas necessitam juntar esforços para criar, na população sob controle, uma base de dados e estrutura familiar apropriada, de forma a permitir a aplicação de metodologias altamente eficientes no uso das informações disponíveis sobre os indivíduos candidatos à seleção, tal como os modelos mistos através do Modelo Animal.
Com base nessa argumentação, o Programa de Melhoramento Genético da Raça Nelore (PMGRN) iniciou, a partir de 1995, a aplicação da Reprodução Programada de Machos Jovens (RPMJ). A idéia básica é introduzir a cada ano, na reprodução sob IA, um número crescente de tourinhos selecionados pelo mérito genético como animais jovens (Diferença Esperada na Progênie – DEPs), no maior número de rebanhos possíveis, com os seguintes objetivos:
a – Multiplicar, da maneira mais rápida, o material genético superior gerado nos rebanhos do PMGRN;
b – Criar uma estrutura familiar e, portanto, dos dados, cada vez mais adequada para a aplicação do Modelo Animal na avaliação genética;
c – Programar os acasalamentos a fim de criar uma distribuição e conexão das informações entre rebanhos, progressivamente mais adequada para obter o máximo proveito dos recursos e técnicas disponíveis;
d – Promover a entrada em centrais de IA de touros à menor idade e com avaliação genética mais acurada, a fim de facilitar sua rápida multiplicação nos rebanhos comerciais, expressão econômica do trabalho genético;
e – Acelerar o progresso genético para toda a raça, propiciando o fluxo do material genético entre os rebanhos integrantes do PMGRN, assim como para o resto da população.
Cabe ressaltar que não se trata de um teste de progênie tradicional, mas sim da reprodução programada dos machos jovens, identificados como geneticamente superiores por meio da avaliação com Modelo Animal usando-se toda a informação disponível, isto é, touros, matrizes e animais jovens conjuntamente. Desta forma, se disporá nas avaliações subseqüentes sobre estes tourinhos, de informações mais completas em termos de acurácia na predição do valor genético: as informações de suas progênies. Tal procedimento permitirá obter predições do valor genético dos touros a uma menor idade e com a máxima acurácia.
Por outro lado, este processo beneficiará em maior grau os rebanhos que disponham de uma parte de suas matrizes para este objetivo, pois tratando-se de tourinhos selecionados pelo Mérito Genético Total (MGT), suas progênies, como média, terão um valor genético alto. Estes produtos passarão a ser parte desses rebanhos com grande antecipação, propiciando que o progresso genético seja acelerado e de custo mais reduzido para tais rebanhos. Note-se que quando os touros sejam definitivamente confirmados como reprodutores de alto potencial genético, suas progênies já estarão prontas para entrar em reprodução nos rebanhos que usaram inicialmente o sêmen dos mesmos.
Deve-se destacar que ao usar um grupo de machos jovens de alto mérito genético, mesmo que as acurácias individuais sejam menores, podemos estar seguros de que a acurácia média poderia ser equivalente à de um reprodutor com grande número de filhos.
Estes fatos devem ser levados em consideração pelos criadores uma vez que, sem dúvida, constituem fatores altamente positivos e determinantes do progresso genético que poderia ser alcançado no futuro. Para este fim seria justificado usar todo ano uma proporção significativa das matrizes de cada rebanho.
Em resumo, este processo contribuirá para incrementar o uso da IA de forma programada na população e, como conseqüência, para a criação de uma estrutura familiar e das informações mais favoráveis para a aplicação da metodologia de modelos mistos (Modelo Animal) na avaliação genética, o que determinará um ganho expressivo no progresso genético a ser alcançado.
________________________________________________________
1Dpto. de Produção Animal, Escola de Veterinária
Universidade Federal de Goiás
